Przemyslaw Prusinkiewicz,
Yvette Wrasmus, Brendan Lane, Lawrence D. Harder and Enrico Coen
(2007)
Evolution and development of inflorescence architectures.(花序構造の進化と発生)
Science Vol. 316, No. 5830, pp. 1452-1456.(2007年6月8日号)
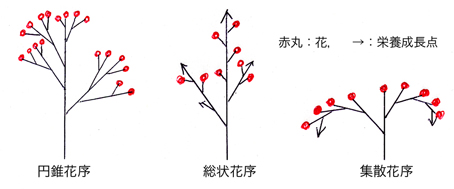
被子植物は,その分類群特有の多様な形態・色の花を持つことを基本形質とするが,その花が単独で茎頂に付く場合(単頂花序という)はむしろ少なく,多くの場合は花の集合体である花序を形成する。この花序は,その構造・形態によって様々な名称が付けられているが,基本となるものは,円錐花序(panicle),総状花序(raceme),および集散花序(cyme)であるとされている(下図参照)。円錐花序は複数回二又分岐した多数の枝(花柄)の先に一斉に花を付けるタイプである。総状花序では,無限成長する主軸(しばしば複数存在)から出る側枝に次々と花を付ける。一方集散花序は,逆に主軸にすぐに花が付き,主軸から形成された側枝が主軸化しまた花を付け,さらにそこから側枝が伸びて主軸化し同様の過程を繰り返すタイプである。実際には,これらの3タイプの複合されたもの,あるいはキク科の頭花のように総状花序の主軸及び側枝が極端に短くなったと考えられているタイプなど,極めて多様な花序の形態が自然界には存在する。しかし進化的・発生的には,以上の3タイプが基本形と考えられており,それからはずれた基本構造はあり得ないとされる。
Prusinkiewiczらによる本論文では,自然界に見られるこの限られた花序の基本型を説明する単純な発生モデルを提案し,更にこのモデルの正しさを,シロイヌナズナを用いた分子遺伝学的実験によって証明した。そのモデルの概要は以下の通りである。(1)花序の構造は,主軸と側枝の頂端分裂組織が栄養成長から生殖成長(花芽形成)へ遷移する時間の違いによって決定される。(2)主軸の分裂組織の遷移時間TAと側枝の遷移時間TBが同じ場合,円錐花序となる。(3)TA>TB(側枝に先に花が付く)の時は総状花序となり,逆に,TA<TB(主軸に先に花が付く)の場合は集散花序となる。
シロイヌナズナは分岐した総状花序を持つ植物であり,その分裂組織の栄養成長状態はTERMINAL FLOWER 1
(TFL1)とLEAFY (LFY)と呼ばれる拮抗する二種類の遺伝子によって調節されている。TFL1は主軸の栄養成長を維持(TAを長くする)し,LFYがそれに対して主軸においてはTFL1の作用を阻害し,また,側枝では頂端分裂組織を栄養成長から生殖成長にシフトさせる(TBを短くする)。そこで,著者らは,この二種類の遺伝子の欠損突然変異体や過剰発現形質転換体を用いることにより,主軸と側枝の頂端分裂組織の栄養成長状態を変化させ,その表現型を分子遺伝学的に解析した。その結果は,彼らのモデルを支持するものであった。
さて,進化的には円錐花序が原始的であり,総状花序や集散花序を持つ植物は,円錐花序を持つ植物から進化したとされている。彼らの報告によると,現生の被子植物では,熱帯多雨林においては円錐花序を持つ植物が有意に多く,又,逆に温帯においては集散花序をもつ植物が有意に多いとされる。また,総状花序をもつ植物も温帯に多い傾向が認められた。円錐花序の場合は,比較的長い栄養成長から生殖成長への遷移時間を経て,一斉に花を付ける。このような性質は,熱帯多雨林のような環境では特に問題はないが,気温や降水量の季節的変化が激しい温帯などにおいては危険を伴う。そのために,生育に不利な季節がある地域に生育分布を広げるにあたっては,長期間にわたって次々と花(および実)を付ける総状花序や集散花序を進化させることが,有利であったと考えられる。一方,総状花序と集散花序間の進化については,主軸と側枝の分裂組織に対して正反対の遺伝的変化を必要とするので,他の成長に関する遺伝子に対する影響も大きく,簡単には行かないであろうと推定している。そのことが,また,多様な花序形態の特定のものがある分類群内で固定される,一つの原因であるのかもしれない。
形態の進化は実験的には証明できないことが多く,理論的研究に終始せざるを得ない場合が多い。本論文では,花序の進化についての理論的考察をシロイヌナズナを用いた実験によって証明するという,少なくとも高等植物においては稀な手法を用いて成功している。今後,このような手法を用いた研究が増えることが予想されよう。